In North America, beech bark disease (BBD) is a complex affecting American beech (F. grandifolia Ehrh.). Its etiology includes the predisposing attack of bark by the beech scale insect, Cryptococcus fagisuga Lind. (Fig. 1), and subsequent invasions and killing of infested bark by several fungi of the genus Nectria (Ehrlich 1934). The principal fungus is N. coccinea var. faginata Lohm. and Watson (Lohman and Watson 1943) (Fig. 2), though N. galligena Bres. also attacks and kills bark predisposed by C. fagisuga (Cotter 1974; Houston 1994a; Mielke et al. 1982). A general framework for its etiology can be expressed as:
Beech trees + C. fagisuga + Nectria spp. => BBD
This correctly implies that a specific chronology of events is required for disease development, and that while the effects of the insect are necessary, the disease is expressed only when Nectria spp. attack scalealtered tissues and, conversely, Nectria attack does not occur unless trees are infested by beech scale.
Brought to Nova Scotia accidentally around 1890 (Ehrlich 1934), C. fagisuga has steadily spread westward and southward through the natural forests of Canada and the United States (Fig. 3). It now is found throughout New England, New York, much of Pennsylvania, northeastern Ohio, northeastern West Virginia, northwestern Virginia, and in a small area of the Great Smokey Mountain National Park on the TennesseeNorth Carolina border (Houston 1994 a, b). Generally, Nectria infections and tree mortality occur 1 to 4 years following a heavy buildup of the insect (Fig. 4) on large trees. The area of current heavy mortality is termed the "killing front" (Fig. 5); regions in which severe mortality occurred earlier comprise the "aftermath zone" (Shigo 1972). In aftermath forests, causal agents are established on small trees of root sprout and seedling origin that often develop in dense stands. Most of the new emerging trees and old survivors become cankered and are rendered highly defective by the scaleNectria complex (Fig. 6).
Biological Factors and Agents Affecting C. Fagisuga
In affected stands, some trees remain free of beech scale and
disease (Fig. 7). Challenge trials have shown them to be resistant
to C. fagisuga (Houston 1982, 1983a). Resistant
trees are found in relatively low numbers (< 1 percent of the
beech stems) and commonly occur in groups (Houston 1983a). The
occurrence of resistant trees in groups is encouraging because
they are easier to recognize than isolated individuals, and are
potentially easier to protect in forest management operations
designed to discriminate against diseased trees. Isozyme genetic
studies have shown that groups of resistant trees originate both
from root sprouts and seed (Houston and Houston 1986, 1990).
Increasing the relative number of resistant trees is important
in reducing the impact of BBD. The results of trials to determine
the effects of various harvesting regimes on the initiation, development,
and survival of root sprouts are being analyzed. In addition,
studies to determine how to clone selected resistant genotypes
have been conducted. Tissueculture techniques in which sprouts
from root segments and forced buds of mature resistant trees are
used have brought several genotypes through to rooting (Barker
et al. 1995). Still needed are trials to develop ways to grow
the tissueculture plantlets in soil and introduce them into the
forest.
Some epiphytes growing on beech bark offer favorable spatial habitats
for C. fagisuga (Ehrlich 1934; Houston et al. 1979).
Infestations often develop initially beneath patches of moss
and lichen. However, not all epiphytes enhance infestations.
In Europe, the common bark fungus Ascodichaena rugosa
sometimes produces a dense, relatively continuous stromatic layer
on European beech, F. sylvatica L. (Butin 1977);
as a consequence, C. fagisuga often is absent on
densely infected bark (Houston et al. 1979). However, trials
initiated in 1975 revealed that although infestation by C.
fagisuga of bark infected by A. rugosa remained
low, stromatic patches sometimes were not sufficiently dense or
complete to preclude significant infestation and subsequent development
of BBD (D. Lonsdale, pers. commun.). In North America, stromatic
patches of A. rugosa often are thin and fractured;
they can offer refuges for C. fagisugasometimes
on trees too small to be infested otherwise (pers. observation).
In Nova Scotia, some stands on steep, southfacing slopes contain
many beech trees that are remarkably free of disease compared
to others in the general area. These trees are heavily colonized
by mosaics of crustose lichens. Several of the predominant lichen
species are rarely colonized by C. fagisuga (Houston
1983b). Such lichens have thalli that are dense, smooth, and
epigenous in contrast to the loosely compact, granularsurfaced
hypogenous thalli of readily colonized species.
To date, no invertebrate parasites of C. fagisuga
have been found, but several predators are known. In North America,
Chilocorus stigma Say. is the most common predator.
C. stigma is most abundant when scale populations
are dense and, although it responds numerically to high scale
densities, its predatory effectiveness is limited by its propensity
to disperse, by its failure to feed on all life stages of scale,
and especially by the high rate of scale reproduction (Mayer and
Allen 1983). Although scale populations on individual trees have
been markedly reduced when populations of coccinellids were high,
their overall effectiveness is limited.
In North America today, scale populations are low in some stands
and regions where they were once high. In some forests, we have
observed precipitous and unexplained population declines. Similar
crashes in other forest insect populations have been associated
with attacks by microbial pathogens.
In England, the entomogenous fungus Verticillium lecanii
Viegas was common where infestations of beech scale were or had
been heavy (Lonsdale 1983). The presence of V. lecanii
depended on high scale density or on coalescense of scale colonies.
It was absent from small, isolated, or new scale colonies because
it spreads from one colony to another by hyphal growth rather
than by aerially dispersed spores (Lonsdale 1983). We do not
know whether V. lecanii or another pathogen is responsible
in North America for observed sharp declines in scale populations
or the maintenance of collapsed populations at low levels.
Agents Affecting Nectria spp.
Nematogonum ferrugineum (Pers.) Hughes (Gonatorrhodiella
highlei) is a biotrophic contact mycoparasite (Barnett
and Binder 1973) that obtains its nutrients from living cells
of its host. The first association of the fungus with BBD was
in North America (Ayers 1941). N. ferrugineum also
was commonly associated with N. galligena both on
cankers of several hardwood species and on beech with BBD (Houston
1983c; Mielke and Houston 1983). The effects of parasitism by
N. ferrugineum in nature are not known even though
high populations of the fungus sometimes occur after severe outbreaks
of BBD.
In culture, growth of parasitized Nectria spp. is little
affected (Blyth 1949; Gain and Barnett 1970); though production
of conidial and perithecial initials is reduced (Shigo 1964).
In inoculation trials, parasitized isolates of N. coccinea
var. faginata and N. galligena spread more
slowly in bark and cambial tissues, and persisted for shorter
periods in bark tissues than unparasitized isolates. Although
cankers resulting from parasitized N. coccinea var.
faginata isolates produced fewer perithecia (Houston 1983c),
this fungus appears ineffective as a biocontrol agent because
in nature it becomes abundant only following severe outbreaks
of BBD.

Figure 1.Beech scale nymph (about 0.3 mm long).

Figure 2.Sexual fruiting bodies (perithecia) of
N. coccinea var. faginata (about 0.3 mm in diameter).
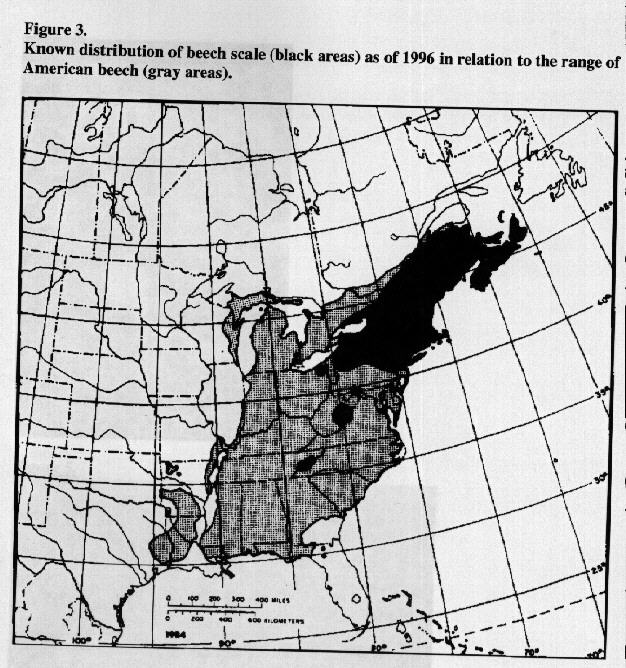
Figure 3.Known distribution of beech scale (black areas) as of
1996 in relation to the range of American beech (gray areas).

Figure 4.Heavy infestations of beech scale
can cover tree boles with white wax.

Figure 5.Tree mortality (trees with bare and discolored crowns)
can be high when forests are affected
by the causal complex for the first time.

Figure 6.Trees in aftermath forests can become
severely defective as cankers accumulate over time.

Figure 7.Some trees are resistant to beech scale
and remain free of disease (left)
in contrast to their susceptible neighbors (right).
 Back to
Back to
SERAMBO
Back to
the Proceedings
Questions?
Email us!